

Using a new HSPC senescence model in vitro to explore the mechanism of cellular memory in aging HSPCs
- PMID: 34365970
- PMCID: PMC8351417
- DOI: 10.1186/s13287-021-02455-x
Using a new HSPC senescence model in vitro to explore the mechanism of cellular memory in aging HSPCs
Retraction in
-
Retraction Note: Using a new HSPC senescence model in vitro to explore the mechanism of cellular memory in aging HSPCs.Stem Cell Res Ther. 2023 Feb 28;14(1):35. doi: 10.1186/s13287-023-03258-y. Stem Cell Res Ther. 2023. PMID: 36855154 Free PMC article. No abstract available.
Abstract
Background: Age-associated changes attenuate human blood system functionality through the aging of hematopoietic stem and progenitor cells (HSPCs), manifested in human populations an increase in myeloproliferative disease and even leukemia; therefore, study on HSPC senescence bears great significance to treat hematopoietic-associated disease. Furthermore, the mechanism of HSPC aging is lacking, especially the cellular memory mechanism. Here, we not only reported a new HSPC senescence model in vitro, but also propose and verify the cellular memory mechanism of HSPC aging of the Polycomb/Trithorax system.
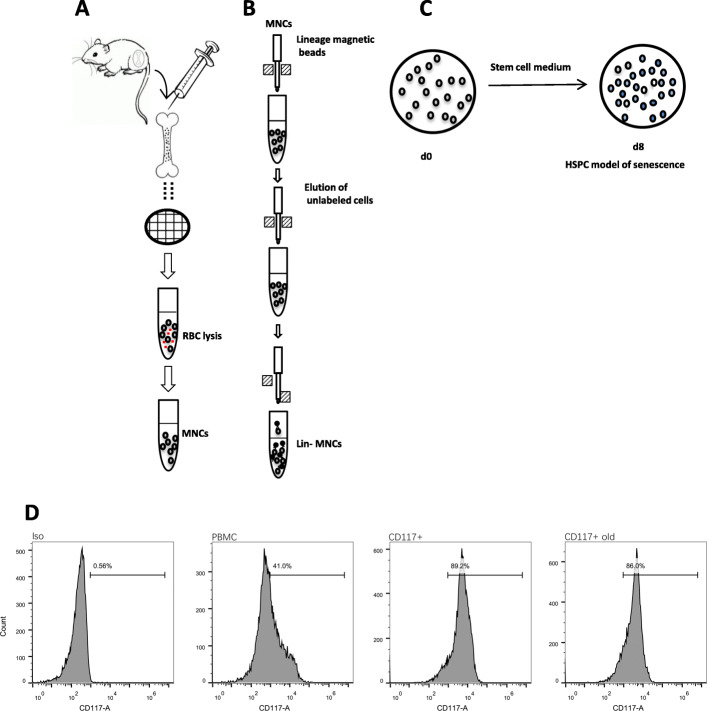
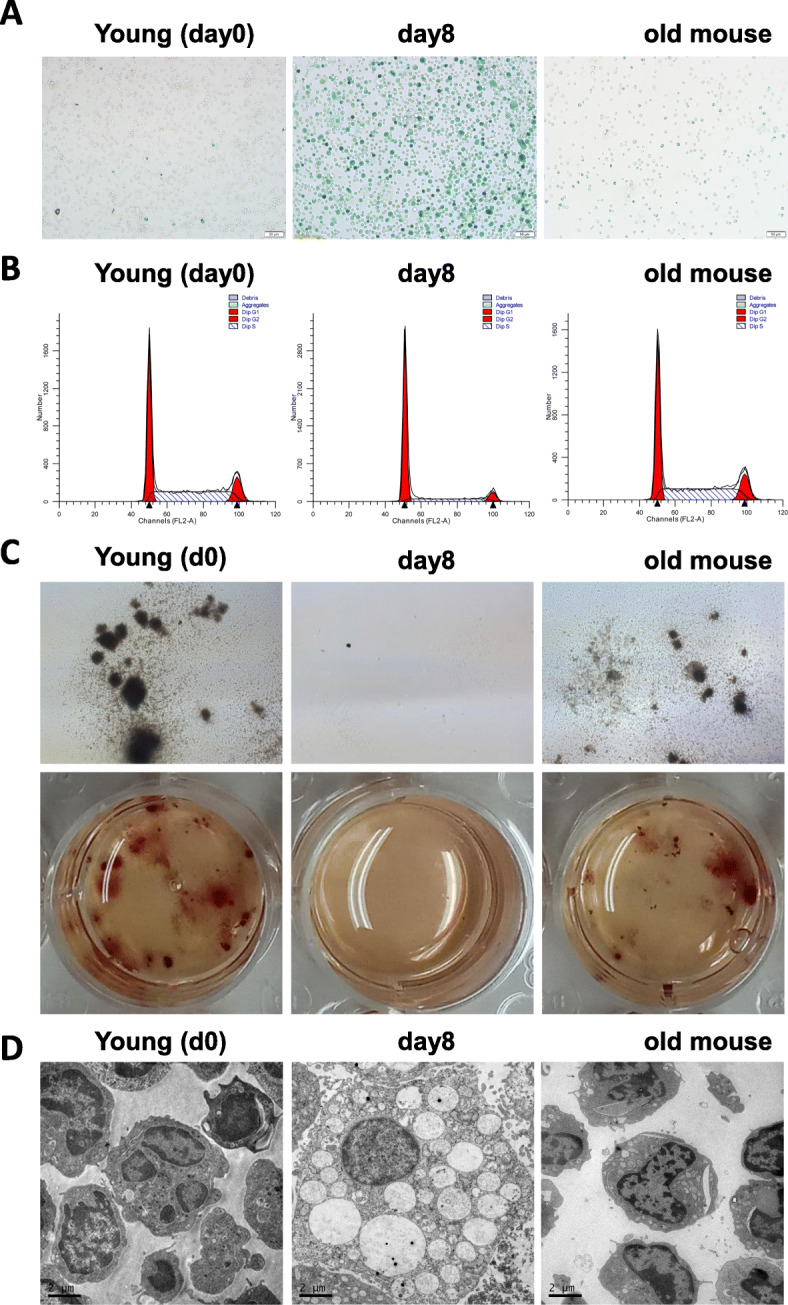
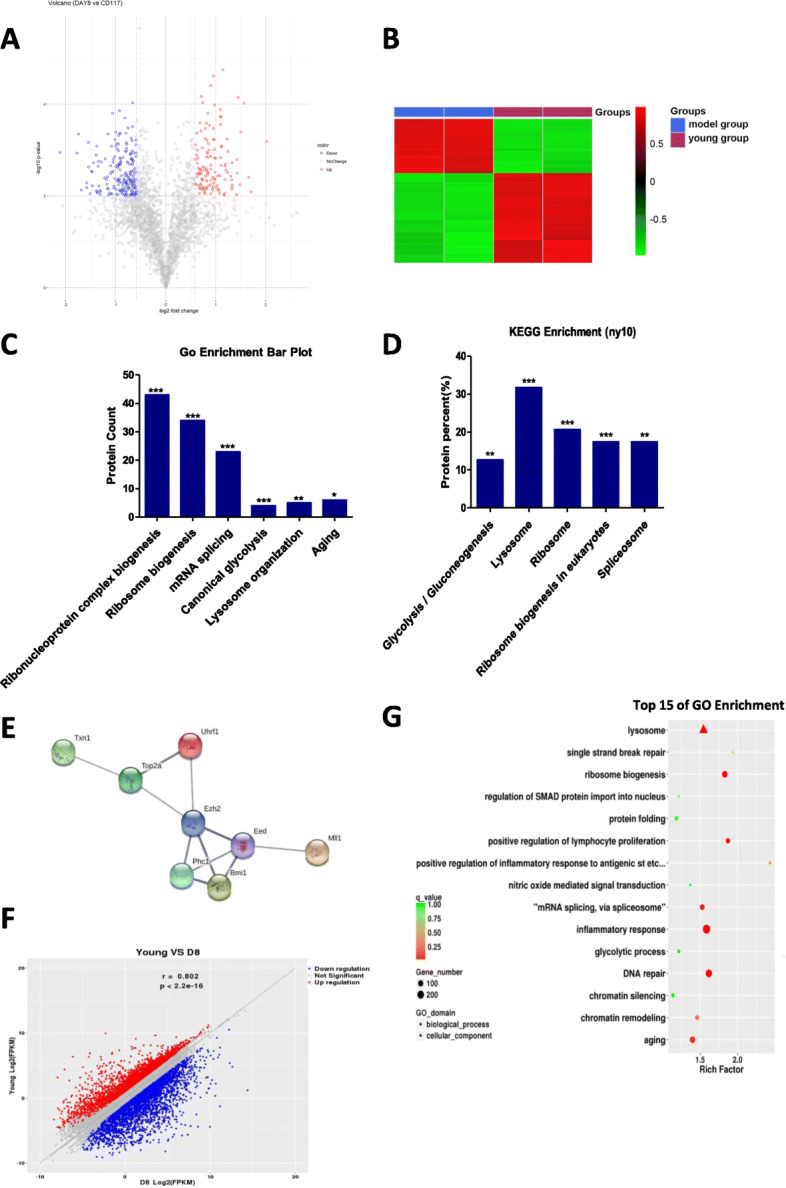
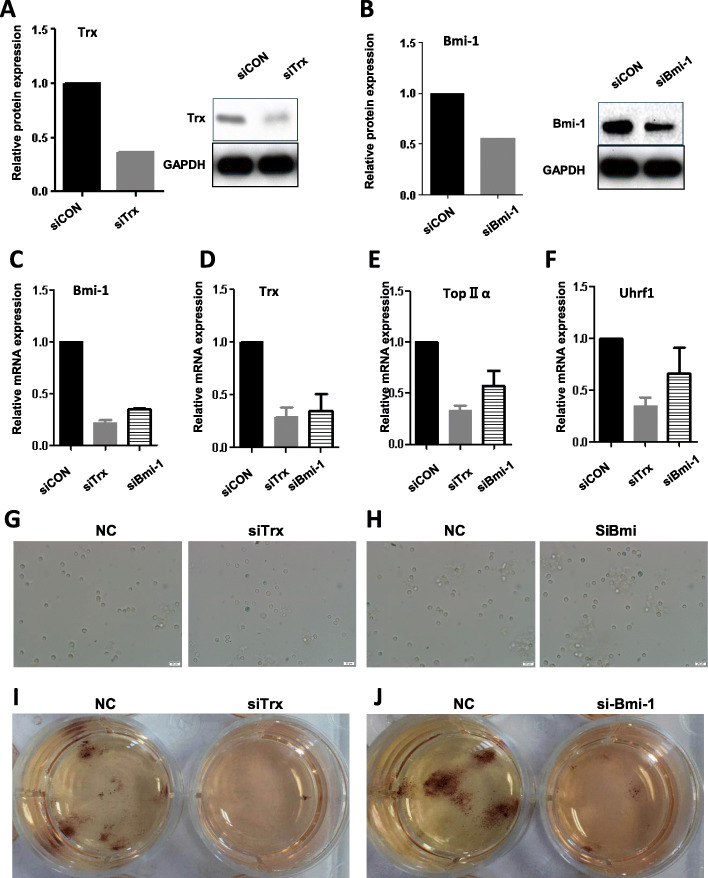
Methods: HSPCs (Lin-c-kit+ cells) were isolated and purified by magnetic cell sorting (MACS). The proportions and cell cycle distribution of cells were determined by flow cytometry; senescence-related β-galactosidase assay, transmission electron microscope (TEM), and colony-forming unit (CFU)-mix assay were detected for identification of the old HSPC model. Proteomic tests and RNA-seq were applied to analyze differential pathways and genes in the model cells. qPCR, Western blot (WB), and chromatin immunoprecipitation PCR (CHIP-PCR) were used to detect the gene expression of cell memory-related proteins. Knockdown of cell memory-related key genes was performed with shRNA interference.
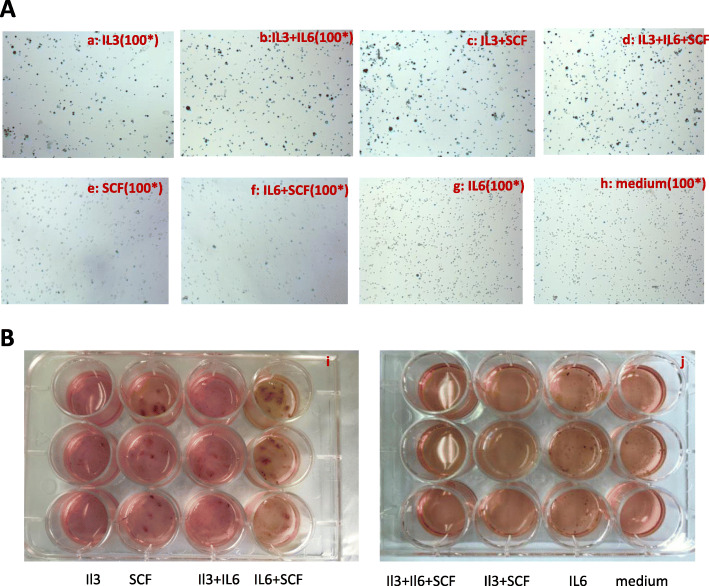
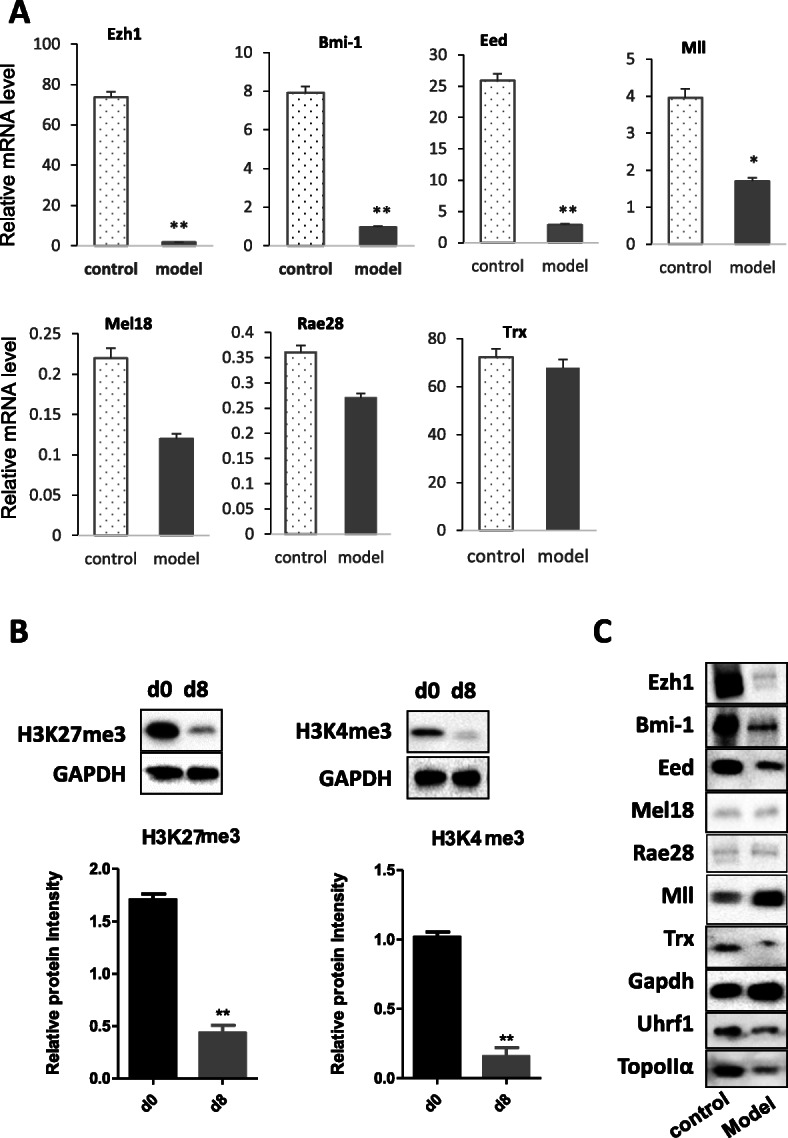
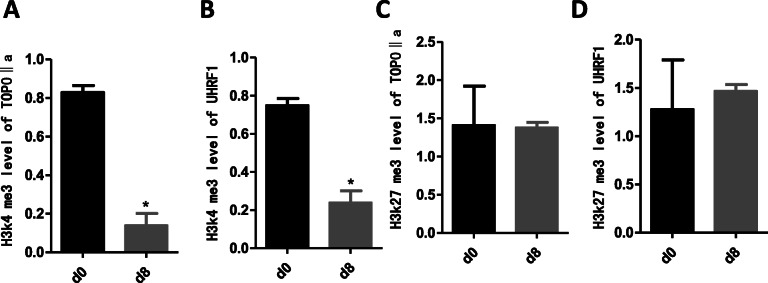
Results: In the model old HSPCs, β-gal activity, cell cycle, colony-forming ability, aging-related cell morphology, and metabolic pathway were significantly changed compared to the young HSPCs. Furthermore, we found the model HSPCs have more obvious aging manifestations than those of natural mice, and IL3 is the major factor contributing to HSPC aging in the model. We also observed dramatic changes in the expression level of PRC/TrxG complexes. After further exploring the downstream molecules of PRC/TrxG complexes, we found that Uhrf1 and TopII played critical roles in HSPC aging based on the HSPC senescence model.
Conclusions: These findings proposed a new HSPC senescence model in vitro which we forecasted could be used to preliminary screen the drugs of the HSPC aging-related hemopathy and suggested cellular memory mechanism of HSPC aging.
Keywords: Cellular memory; HSPC; Senescence model; TOPOIIα; UHRF1.
© 2021. The Author(s).
Conflict of interest statement
The authors declare that they have no competing interests.
Figures
Similar articles
-
Hematopoietic stem/progenitor cell senescence is associated with altered expression profiles of cellular memory-involved gene.Biosci Rep. 2018 Feb 21;38(1):BSR20171589. doi: 10.1042/BSR20171589. Print 2018 Feb 28. Biosci Rep. 2018. PMID: 29363553 Free PMC article.
-
Isobaric tags for relative and absolute quantitation (iTRAQ)-based proteomic analysis of mRNA splicing relevant proteins in aging HSPCs.Aging Clin Exp Res. 2021 Nov;33(11):3123-3134. doi: 10.1007/s40520-020-01509-z. Epub 2020 Mar 5. Aging Clin Exp Res. 2021. PMID: 32141009
-
An early-senescence state in aged mesenchymal stromal cells contributes to hematopoietic stem and progenitor cell clonogenic impairment through the activation of a pro-inflammatory program.Aging Cell. 2019 Jun;18(3):e12933. doi: 10.1111/acel.12933. Epub 2019 Mar 3. Aging Cell. 2019. PMID: 30828977 Free PMC article.
-
Glucose Metabolism and Aging of Hematopoietic Stem and Progenitor Cells.Int J Mol Sci. 2022 Mar 11;23(6):3028. doi: 10.3390/ijms23063028. Int J Mol Sci. 2022. PMID: 35328449 Free PMC article. Review.
-
Advances in umbilical cord blood stem cell expansion and clinical translation.Exp Hematol. 2015 Jul;43(7):498-513. doi: 10.1016/j.exphem.2015.04.011. Epub 2015 May 10. Exp Hematol. 2015. PMID: 25970610 Review.
References
-
- Li J, Cai D, Yao X, Zhang Y, Chen L, Jing P, et al. Protective effect of Ginsenoside Rg1 on hematopoietic stem/progenitor cells through attenuating oxidative stress and the Wnt/beta-catenin signaling pathway in a mouse model of d-galactose-induced aging. Int J Mol Sci. 2016;17(6):849. - PMC - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
